- Home
- News
- Spotlight on Science
- Structure of the...
Structure of the human ferritin-transferrin receptor 1 complex
12-04-2019
The human transferrin receptor 1 (CD71) is a transmembrane protein responsible for iron uptake in cells. The cryo-EM structure of the CD71 ectodomain bound to human ferritin (H-Ft) reveals that H-Ft binds to a CD71 region different to that recognised by transferrin. However, it significantly overlaps with a region of CD71 recognised by certain pathogens prior to cell invasion.
The transferrin receptor 1 (CD71) plays a pivotal role in iron uptake in all vertebrates and its function is crucial for the regulation of iron homeostasis and for cell viability. To import iron into cells, CD71 binds iron-loaded transferrin (Tf) and undergoes constitutive internalisation by activation of the clathrin-dependent endocytic machinery. CD71 has been also shown to mediate the uptake of heavy chain ferritin (H-Ft) from serum as an alternative or additional source of bioavailable iron [1]. Thanks to a combination of its cage-like structure, its unique cargo capabilities for small therapeutic molecules or diagnostic agents and its recognition selectivity towards CD71 [2], H-Ft has recently moved to centre stage as a privileged protein carrier for drug/diagnostics encapsulation and drug-delivery. Indeed, since the receptor is highly expressed in most cancer cell types, the H-Ft/CD71 system could represent a very convenient nano-tool for selective delivery of therapeutic and diagnostic payloads to tumour tissues. Additionally, CD71 is also a preferred entry carrier for human pathogenic arenaviruses [3, 4] and hepatitis C virus [5] as well as feline and canine specific parvoviruses [6]. Recently, Plasmodium vivax, the most common malaria parasite, was found to access reticulocyte cytoplasm by recognising the same CD71 epitope as arenaviruses [7].
CD71 is a homodimeric type II transmembrane protein composed of a small cytoplasmic domain, a single-pass transmembrane region, and a complex extracellular ectodomain. The crystal structure of the human transferrin receptor-transferrin complex [8] shows the ectodomain to comprise three regions: a protease-like domain in contact with the cell membrane, a helical domain comprising the dimer contact regions, and an apical domain (Figure 1a). Transferrin is harboured between the membrane and the basal portion of the ectodomain contacting only its helical and protease-like domains (Figure 1b). The apical domain does not participate in receptor-transferrin interactions and freely displays its binding epitopes recognised by viral and plasmodium proteins (Figure 1b). A key missing piece of information in this picture, however, are the structural determinants of the interaction between CD71 and H-Ft.
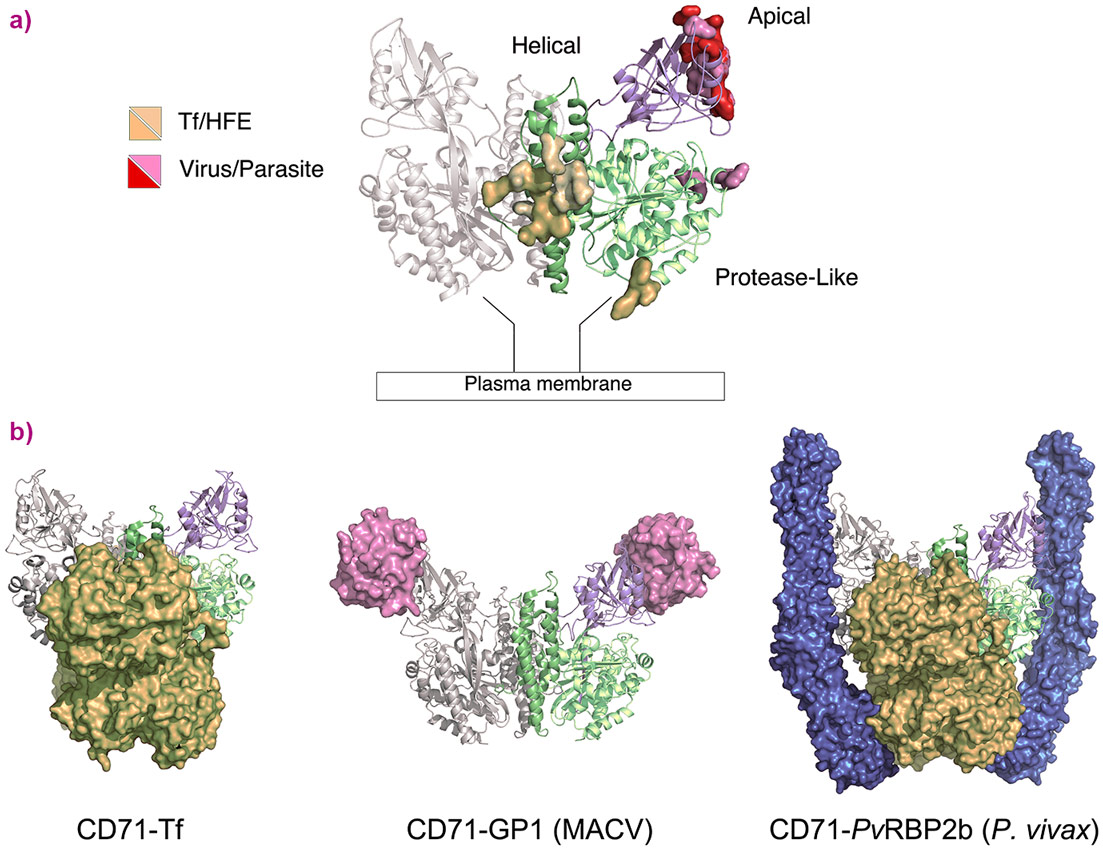 |
|
Figure 1. CD71 receptor: ligand recognition epitopes and binding modes. CD71 homodimer is represented as ribbon (pdb 3KAS [4]). One monomer is in light grey, the other is coloured variably to the receptor domains (apical, violet; protease-like, lime; helical, green). a) CD71 recognition epitopes for Tf/HFE and viruses/parasite are represented as orange/wheat and red/pink surfaces, respectively. b) CD71 is shown in complex with Tf (orange surface, pdb 1SUV [9]), with the GP1 protein of MACV (pink surface, pdb 3KAS), and with Tf (orange surface) and PvRBP2b from P. vivax (blue surface, pdb 6D04 [7]). |
In this work, cryo-electron microscopy data collected at the Titan Krios microscope at CM01 allowed the reconstruction by single particle analysis of a map of the H-Ft/CD71 complex at 3.9 Å resolution (Figure 2a). H-Ft binds CD71 at the apical domain with contacting regions including the βII-1, βII-2 and βII-8 strands and αII-2 helix of the receptor domain (Figure 2b). This specific portion of CD71 does not overlap with the transferrin binding region of CD71. Instead, it largely corresponds to the central epitope of CD71 which determines the binding and subsequent cellular entry of viruses and the malaria parasite. In particular, three amino acids (Y211, L212 and N215 on the βII-2 strand) are common contacts for all the CD71 apical domain ligands, i.e. H-Ft, different viruses and P. vivax proteins. Mutations on H-Ft amino acids contacting the common residues of CD71 strongly affect ferritin-receptor interactions, as observed by surface plasmon resonance binding assays and by cellular ferritin uptake experiments.
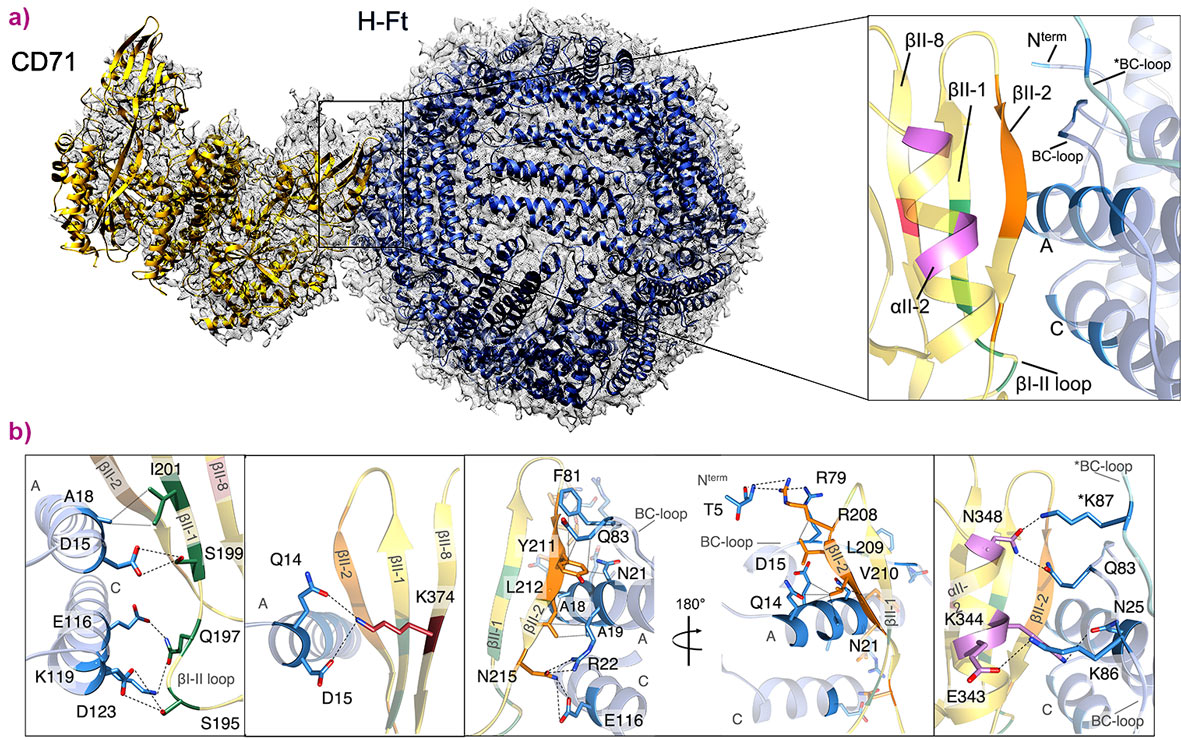 |
|
Figure 2. Structure of the human CD71/H-Ft complex. a) Atomic model of the complex of human CD71 (yellow) and human H-Ft (blue) shown fitted in the cryo-EM map at a global resolution of 3.9 Å (grey mesh). On the right, close-up view of the contact region. The interacting residues of CD71 are highlighted by different colours depending on the specific secondary structural element of the apical domain to which they belong. Interacting residues of H-Ft are colored in blue. b) CD71 and H-Ft interacting regions. Contacting residues within 5 Å distance are shown in sticks, labelled and coloured according to the colour code used for secondary structure in panel (a) (right side). Dashed black lines indicate electrostatic interactions. Dotted grey lines represent hydrophobic contacts. |
The structure of the H-Ft/CD71 complex revealed the specific sites on CD71 exploited by ferritin for physiological access to the cell through CD71. Even a single subunit of H-chain within the 24-mer of ferritin, can be recognised by CD71 and this might be sufficient to favour the cellular internalisation of the mixed H/L-chain 24-mer ferritin through this route of access, even though in serum the L-chain is prevalent in the 24-meric assembly. Moreover, it accounts for the phenomenon of transferrin-independent binding of ferritin to the receptor, allowing differential regulation of iron uptake, and indicates a previously unsuspected physiological role for the CD71 apical domain. Importantly, pathogens have adapted to exploit the ferritin epitope to unlock cellular barriers by mimicking this physiological interaction with the CD71 apical domain and the ferritin-binding function now demonstrated for this domain might prevent an “arms race” by hosts to prevent cell entry [3].
Finally, this work provides a sound structural basis for the development of ferritin-like anti-viral or anti-parasite therapeutic ligands, such as antibodies or peptidomimetics, capable of blocking the “common contacts” epitope on CD71 residue, and to further engineering of ferritins as nanocarriers and theranostic agents.
Principal publication and authors
Cryo-EM structure of the human ferritin–transferrin receptor 1 complex, L.C. Montemiglio (a,b,c), C. Testi (a,d), P. Ceci (b), E. Falvo (b), M. Pitea (a), C. Savino (b), A. Arcovito (e,f), G. Peruzzi (d), P. Baiocco (d), F. Mancia (g), A. Boffi (a), A. des Georges (h,i,j), B. Vallone (a,c), Nat. Commun. 10 (1):1121. doi: 10.1038/s41467-019-09098-w.
(a) Dept of Biochemical Sciences “Alessandro Rossi Fanelli”, Sapienza University of Rome (Italy) (b) Institute of Molecular Biology and Pathology, National Research Council, Rome (Italy)
(c) Istituto Pasteur-Fondazione Cenci Bolognetti, Dip. di Scienze Biochimiche “A. Rossi Fanelli”, Sapienza Università di Roma (Italy)
(d) Center for Life Nano Science @ Sapienza, Istituto Italiano di Tecnologia, Rome (Italy)
(e) Istituto di Biochimica e Biochimica Clinica, Università Cattolica del Sacro Cuore, Rome (Italy)
(f) Fondazione Policlinico Universitario Agostino Gemelli-IRCCS, Rome (Italy)
(g) Dept. of Physiology and Cellular Biophysics, Columbia University Medical Center, New York (USA)
(h) Advanced Science Research Center at The Graduate Center of the City University of New York (USA)
(i) Dept. of Chemistry and Biochemistry, City College of New York (USA)
(j) Programs in Biochemistry and Chemistry, The Graduate Center of the City University of New York (USA)
References
[1] L. Li, et al., Proc Natl Acad Sci U S A 107, 3505-3510 (2010).
[2] M. Truffi, et al., Pharmacol Res 107, 57-65 (2016).
[3] J. Abraham, et al., Nat Struct Mol Biol 17, 438–444 (2010).
[4] A. Demogines, et al., PLoS Biol 11, e1001571 (2013).
[5] S.R. Radoshitzky, et al., Plos One 6, e21398 (2011).
[6] D.N. Martin, et al., Proc Natl Acad Sci U S A 110, 10777-10782 (2013).
[7] S. Hafenstein, et al., Proc Natl Acad Sci U S A 104, 6585-6589 (2007).
[8] J. Gruszczyk, et al., Science 359, 48-55 (2018).
[9] C.M. Lawrence, et al., Science 286, 779-782 (1999).
[10] Y. Cheng, et al., Cell 116, 565-576 (2004).
partners



European Synchrotron Radiation Facility - 71, avenue des Martyrs, CS 40220, 38043 Grenoble Cedex 9, France.