- Home
- News
- General News
- Joining back together...
Joining back together the ends of broken DNA
16-03-2017
The DNA in our genes is constantly under attack from chemicals and radiation. Broken DNA is repaired by a collection of proteins including the enzyme called DNA-PKcs. The crystal structure of DNA-PKcs, solved following data collection at the ESRF, gives valuable insight into the start of the DNA repair process and hints at a selection mechanism for different repair paths.
Share
Chemicals from metabolic activities and environmental factors such as radiation cause thousands of lesions per day in the genetic information stored in the chromosomes inside each of our cells. These lesions, if uncorrected, could prevent the genes from being read during the production of proteins. The damage could even result in changes to the code, which in the worst case could lead to mutations in daughter cells and eventually cancer. Fortunately, each cell contains enzymes whose purpose is to maintain the chromosomes by repairing any damage to the DNA, or else to signal the start of programmed cell death to prevent severely damaged genes from being passed on to daughter cells.
DNA maintenance is carried out by many different enzymes and proteins working together to search for and repair damaged DNA. Two principal repair mechanisms are known for double strand breaks. If there is a sister chromatid available to provide a template, so called “homologous recombination” is used to ensure correct reconstruction of the gene sequence. Otherwise the broken ends are joined back together directly, sometimes involving errors in the genome sequence; this pathway is called “non-homologous end joining”.
Lynn Sibanda, Dima Chirgadze and David Ascher working with Sir Tom Blundell at the University of Cambridge, UK, studied a key enzyme involved in DNA repair, the DNA-dependent protein kinase catalytic subunit (DNA-PKcs). Following data collection at ESRF beamlines ID23-1 and ID29, they obtained the crystal structure of human DNA-PKcs in complex with the C-terminal peptide of Ku80 with a resolution of 4.3 Å, which contained sufficient detail to advance our knowledge of these two proteins for the initiation of the DNA repair process, gaining in detail over earlier X-ray and cryo-EM studies of DNA-PKcs.
Blundell described key steps leading to the interpretation of this enzyme complex, “This is the longest single chain globular protein whose structure has been defined by X-ray crystallography. It has over eighty repeated structures, known as HEAT repeats, containing helix-turn-helix motifs, which made following the registration of the sequence in the electron density very challenging. We used anomalous scattering from seleno-methionines to monitor this – it could only be done well at a synchrotron”.
 |
|
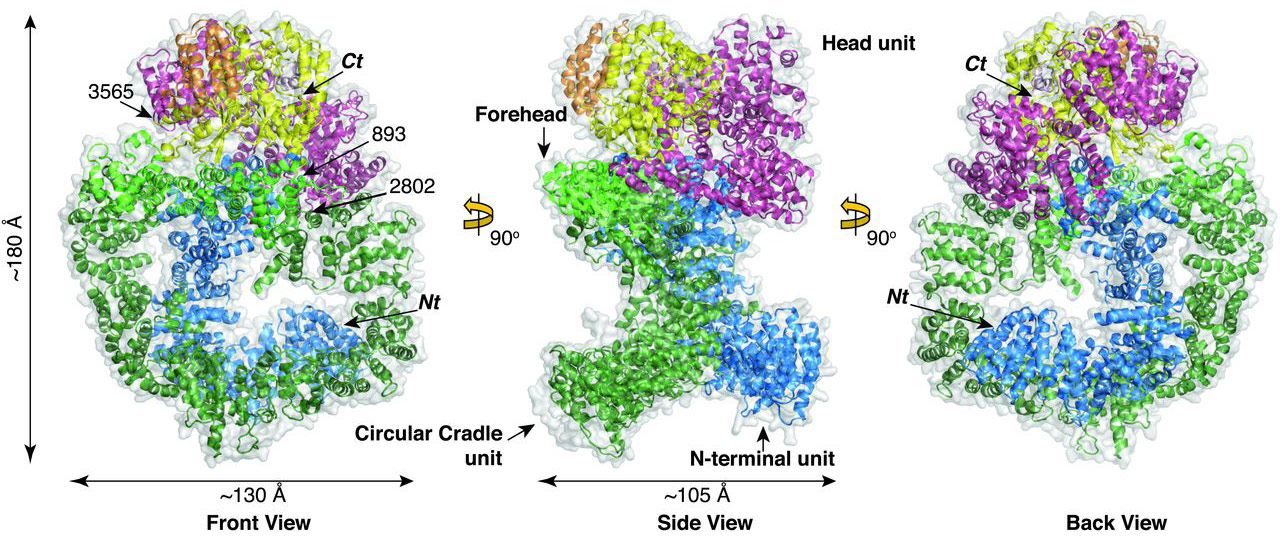
Overall crystal structure of DNA-PKcs. This 4128–amino acid structure comprises three large structural units: N-terminal unit (Nt), Circular Cradle, and Head. |
As expected, the structure of DNA-PKcs kinase region was found to be similar to other kinases, containing an activation loop as well as the catalytic loop of the active site. Access to the active site is thought to require substantial conformational changes in many parts of the protein. Differences between the two copies of DNA-PKcs in the crystal structure (the asymmetric unit contained two molecules) provided evidence of the extent of these changes, the purpose of which could be to ensure selectivity of the enzyme towards its substrate through an allosteric mechanism. Hence, the binding of DNA-PKcs to Ku80 is believed to cause a change in DNA-PKcs that opens the pathway to the active site, effectively activating the enzyme.
The scientists also observed that the putative BRCA1 binding site is near to the Ku80 binding site (BRCA1 is a protein involved in the template-based repair mechanism). This led to the hypothesis that BRCA1 may influence the binding of Ku80 to DNA-PKcs, providing a mechanism to switch between the pathways of the two repair mechanisms, with BRCA1 activating the template-based pathway.
For the end-joining pathway, the dimeric arrangement of the two DNA-PKcs molecules observed in the crystal structure may facilitate the bringing together of each end of the double-strand break.
 |
|
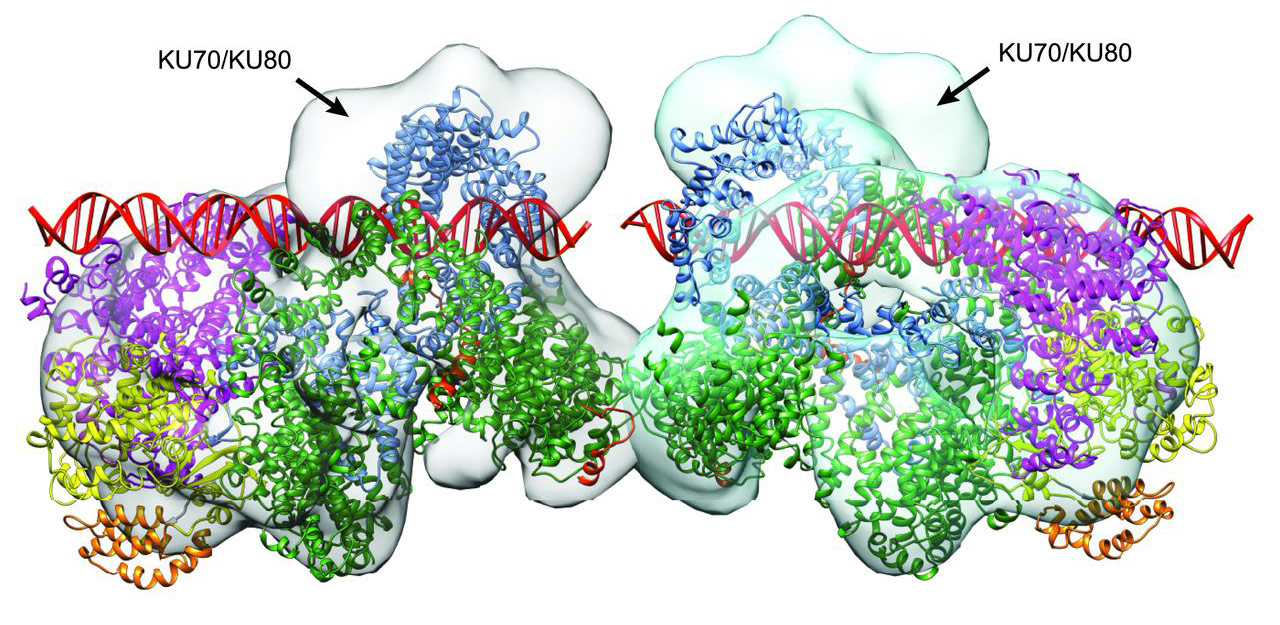
Model of DNA-PKcs and Ku70/80 showing position of DNA. Model of the two DNA-PKcs molecules in the crystallographic asymmetric unit showing possible positions for the DNA helices (in red) bound to each DNA-PKcs, overlaid on a cryo-EM map of DNA-PKcs bound to Ku70/80, showing the positions of the Ku complexes relative to the overall structure. |
The sequence of the DNA repair is now believed to occur as follows: a Ku80 containing protein complex (Ku70/Ku80 heterodimer) binds to the broken ends of DNA and recruits DNA-PKcs, switching on this enzyme via the conformational change that opens its active site. The active DNA-PKcs can then autophosphorylate itself and phosphorylate other proteins, effectively raising a flag to signal the damage to other enzymes. Competition between BRCA1 and Ku80 causes selection of one of the two repair pathways, with Ku80 holding DNA-PKcs on the end-joining pathway. Other enzymes (Artemis/XRCC4/XLF/, etc.) then take their turn to organise in space and time the repair of the break by DNA ligase IV, before the KU complex and DNA-PKcs are released.
This research helps us to understand the functioning of an intricate process within the human body that could be key for future pharmaceuticals development. Blundell explains, “We will use the structure to guide targeting of this system with new drugs. DNA repair often becomes uncontrolled in cancer and its inhibition would help to kill tumours”.
Publication
DNA-PKcs structure suggests an allosteric mechanism modulating DNA double-strand break repair B.L. Sibanda, D.Y. Chirgadze, D.B. Ascher, T.L. Blundell, Science 355, 520–524 (2017); doi: 10.1126/science.aak9654.
Text by Gary Admans; Images reprinted from B.L. Sibanda et al., Science 355, 520–524 (2017), Reprinted with permission from AAAS.
partners



European Synchrotron Radiation Facility - 71, avenue des Martyrs, CS 40220, 38043 Grenoble Cedex 9, France.