- Home
- Ser or Leu: Structural impact of a genetic code alteration in C. albicans
Ser or Leu: Structural impact of a genetic code alteration in C. albicans
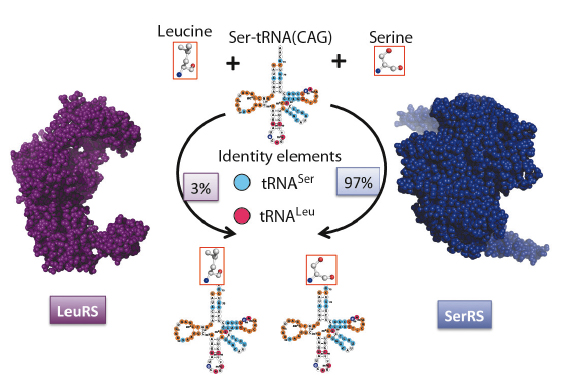
A 200-million year old alteration to the standard genetic code resulted in the appearance of a novel serine tRNA [1], allowing the human pathogen Candida albicans and other CTG-clade fungi to translate the otherwise universal leucine CUG codon as serine. Due to a mutation in the anticodon loop, this unusual serine tRNA recognises the CUG codon, but additional changes in its sequence allow recognation both by the seryl- and the leucyl-tRNA synthetases (Figure 97). Therefore, the serine-tRNACAG can be aminoacylated either with serine (97%) or leucine (3%), so that proteins encoded by a CUG-containing gene incorporate either of these amino acids at the CUG-encoded positions, with a significant bias towards serine. This ambiguous CUG decoding affects approximately 67% of the C. albicans ORFeome and leads to proteome expansion. Curiously, C. albicans can be engineered to increase the percentage of leucine misincorporation up to 28%, leading to impressive phenotypic alterations. However, these show no changes in growth rates, implying that this pathogenic yeast has the ability to survive with significant CUG ambiguity.
This Candida Spp. serine/leucine ambiguity generates an intriguing question from a structural biology perspective: how can the 4168 CUG-encoded C. albicans proteins cope with incorporation of serine or leucine at the same position without significant protein misfolding and consequent cell dysfunction? Previous data showed that CUG codon ambiguity in CTG-clade fungi resulted in the relocation of the CUG codons to novel positions: 98% of the ancestral CUGs disappeared and were replaced by other leucine codons (UAA, UAG) and new CUGs were located in positions occupied by codons coding for serine or other polar amino acids in homologous genes from related fungal species. A large-scale analysis of the location of CUG-encoded residues in CTG-clade proteins showed that they reside predominately in non-conserved regions of the sequence, avoiding the buried protein core. Consequently, it is expected that the structure and function of protein isoforms with serine or leucine at CUG positions does not differ significantly and that increased incorporation of leucines at these positions does not result in massive protein misfolding.
Nevertheless, the substitution of serine for leucine (and vice versa) in CUG-containing genes is unlikely to be completely neutral, since surface residues are often involved in macromolecular interactions, which might be compromised upon modification of the molecular surface. Moreover, structural and functional changes can be predicted for a small number of proteins containing CUG-encoded residues in functionally conserved positions, possibly explaining the phenotypic and morphological changes accompanying increased leucine incorporation in C. albicans [2].
 |
|
Fig. 97: The molecular determinants of genetic code ambiguity in Candida spp. C. albicans serine-tRNACAG is a hybrid molecule with specific identity elements, which allow the recognition both by seryl- and leucyl-tRNA synthetases. This tRNA is predominantly charged with serine (97%) highlighting the predominant role of SerRS in ambiguous CUG decoding. |
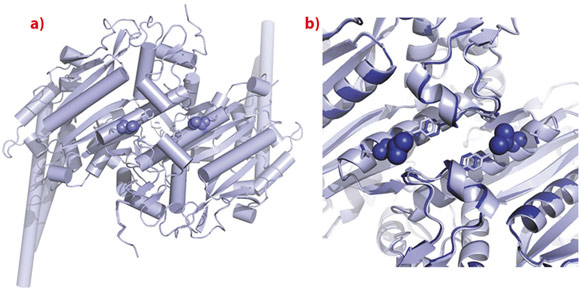
The seryl-tRNA synthetase (SerRS) is the main player in CUG-decoding ambiguity in C. albicans, and itself contains a CUG-encoded residue in a position where polar amino acids are preferred in homologous yeast SerRSs. The three-dimensional structures of the two isoforms of the enzyme were determined, showing that no large structural change results from serine replacement by leucine at the CUG position. The CUG-encoded residue is located at the dimer interface, partially exposed at the protein surface, and it is not involved in inter-monomer interactions (Figure 98). Nonetheless, the presence of the bulkier hydrophobic leucine side chain induces local rearrangements in the adjacent monomer in the vicinity of the C-terminal tail, unique in cytoplasmic eukaryotic SerRSs and required for enzyme stability and function [3]. These small structural changes are associated with functional differences between the two isoforms, indicating that although the placement of serine or leucine at the CUG-encoded position does not induce major structural changes or protein misfolding, the two isoforms have small but significant differences in function.
 |
|
Fig. 98: Crystal structures of C. albicans seryl-tRNA synthetase isoforms. a) Structure of the SerRS_Ser197 dimer with Ser197 shown as spheres. b) Local structural changes at the CUG-encoded position in the SerRS_Leu197 isoform. |
Similar functional alterations could be expected for the fungal proteins containing CUG-encoded residues in functionally relevant positions, namely those playing a role in pathogenesis-associated signalling cascades. Therefore, besides providing a deeper insight into the molecular basis of CUG codon relocation in the CTG clade fungi and highlighting the intrinsic link between natural codon ambiguity and protein multifunctionality, our results also provide new perspectives for future research in Candida virulence pathways.
Principal publication and authors
R. Rocha (a), P.J. Barbosa Pereira (a), M.A. Santos (b) and S. Macedo-Ribeiro (a), Proc Natl Acad Sci USA. 108, 14091-6 (2011).
(a) IBMC – Instituto de Biologia Molecular e Celular, Porto (Portugal)
(b) CESAM and Universidade de Aveiro, Aveiro (Portugal)
References
[1] M.A. Santos, G. Keith and M.F. Tuite, Embo J 12, 607-616 (1993).
[2] I. Miranda, R. Rocha, M.C. Santos, D.D. Mateus, G.R. Moura, L. Carreto and M.A. Santos, PLoS ONE 2, e996 (2007).
[3] M. Mocibob and I. Weygand-Durasevic, Arch Biochem Biophys 470, 129-138 (2008).
partners



European Synchrotron Radiation Facility - 71, avenue des Martyrs, CS 40220, 38043 Grenoble Cedex 9, France.