- Home
- Towards atomic resolution of the plant photosystem I supercomplex
Towards atomic resolution of the plant photosystem I supercomplex
The conversion of solar energy into chemical bonds by the photosynthetic machinery of plants, green algae and cyanobacteria is essential for the survival of all higher life forms. The primary event of this process - light induced charge translocation - is catalysed by photosystem I (PSI) and photosystem II (PSII), two multisubunit protein complexes embedded in the thylakoid membrane and acting in series. PSI harnesses light energy to transfer an electron from the soluble copper protein, plastocyanin, at one side of the membrane to ferredoxin and eventually NADPH on the opposite side. In the dark, NADPH (and ATP) provides the energy for assimilation of CO2 into carbohydrates. In 2003, the structure of plant PSI was solved at 4.4 Å resolution [1]. The components visible in this crystal structure were a β-carbon model of 16 proteins, including 45 transmembrane helices, 2 phyloquinones, 3 Fe4S4 clusters and 167 chlorophyll molecules.
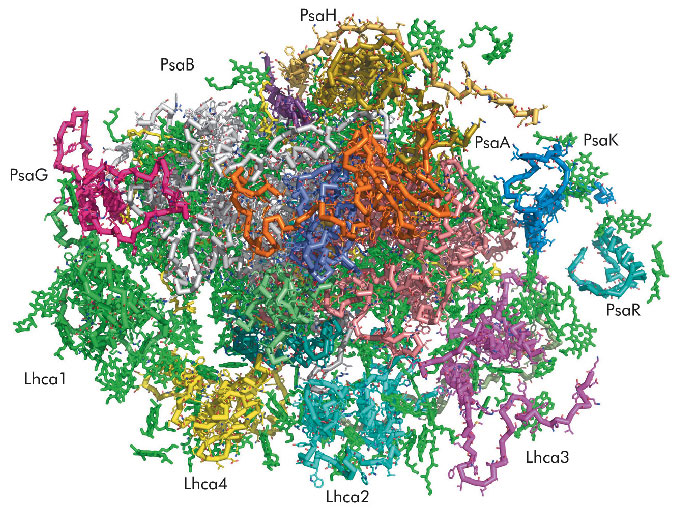
Continuous and progressive improvement of crystal quality and the ready access to ESRF resulting from the BAG programme have enabled recent determinations of the crystal structure of PSI first to 3.4 Å [2] then to 3.3 Å resolution. One of the most important factors that allowed higher resolution data to be obtained was the availability of the micro-focussing MX end station ID23-2. We were among the first to realise the advantage of using a highly-focussed beam with relatively large samples. For PSI, average crystal dimensions are 100x30x5 µm3 and, because they are so thin, they have a tendency to bend. This tendency results in elongated and much less clear spots in diffraction patterns taken with larger X-ray beams. The high intensity micro-focussed beam used together with a recently developed helical-motion data collection capability resulted in much better data sets that revealed several components of PSI that had not been detected in previous crystal structures (Figure 100).
 |
|
Fig. 100: A view from the stroma of the at 3.3 Å crystal structure of plant PSI. The 14 subunits of the reaction centre and the four light-harvesting proteins (Lhca1 - Lhca4) are depicted in different colours. The chlorophylls and other prosthetic groups are as described in Figure 101. |
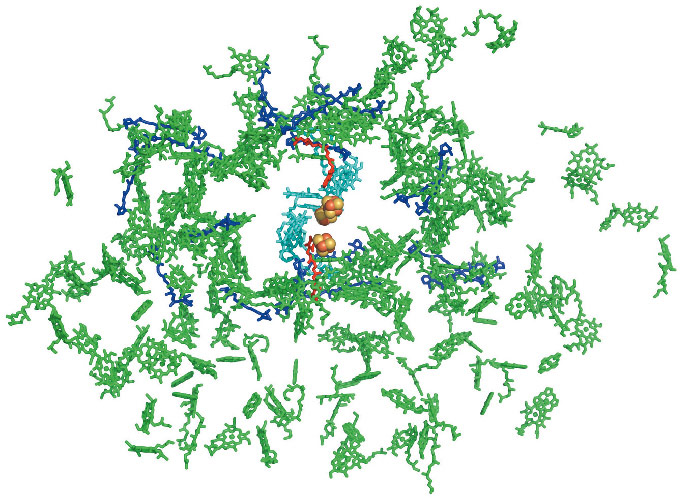
The current 3.3 Å resolution plant PSI crystal structure reveals 18 protein subunits containing 46 transmembrane helices, 173 chlorophylls, 18 β-carotenoids, 3 Fe4S4 clusters and 2 phyloquinones (Figure 101). Ten of the 12 plant PSI reaction centre core complex subunits (PsaA, PsaB, PsaC, PsaD, PsaE, PsaF, PsaI, PsaJ, PsaK, and PsaL) are closely related to those of the PSI of cyanobacteria. Other subunits (PsaG and PsaH) have unique structural functions related to interactions of the reaction centre core with four light-harvesting complexes of LHCI (Lhca1 - Lhca4) and provide a potential site of interaction with LHCII. The four LHCI proteins assemble into two dimers arranged in series and form a half-moon shaped belt that docks to the subunit F side of the reaction centre. The association of the PSI core with the LHCI belt is both weak and asymmetric (Figure 100). Only one of the four Lhca proteins (Lhca1) is firmly attached to the reaction centre. It may serve as an “anchor” for the assembly of the other Lhca monomers. Formation of the LHCI dimers and the interactions between them does not involve transmembrane helices. However, the solvent exposed C and N termini of each Lhca monomer attach to solvent exposed loops on the neighbouring monomer.
 |
|
Fig. 101: A view from the stroma on the prosthetic groups currently modelled in the structure of the plant PSI supercomplex. Chlorophyll moieties are coloured in green or, in the case of those involved in electron transfer, cyan. The electron density of 109 of the chlorophyll molecules modelled exhibited complete or partial phytol chains thus allowing assignment of the orientation of their QX and QY transition dipole moments. The 2 phyloquinones modelled are shown in red and the 18 carotenoids moieties in blue. The 3 Fe4S4 clusters are depicted as yellow and brown spheres. |
The structural information on proteins, co-factors and the interactions between them described in this work provides a significant step towards our understanding of how the very large PSI supercomplex has evolved. Nevertheless, only atomic resolution structures of the two components of PSI - the reaction centre and the LHCI - will allow calculations designed to explain the unprecedented quantum yield of this PSI system. We hope that this daunting goal will be achieved before too long.
Principal publication and authors
A. Amunts, H. Toporik, A. Borovikova and N. Nelson, J Biol Chem. 285, 3478-3486 (2010).
Department of Biochemistry, George S. Wise Faculty of Life Sciences, Tel Aviv University (Israel)
References
[1] A. Ben-Shem, F. Frolow and N. Nelson, Nature 426, 630-635 (2003).
[2] A. Amunts, O. Drory and N. Nelson, Nature 447, 58-63 (2007).
partners



European Synchrotron Radiation Facility - 71, avenue des Martyrs, CS 40220, 38043 Grenoble Cedex 9, France.