- Home
- Architecture of respiratory complex I: the “steam engine” of the cell?
Architecture of respiratory complex I: the “steam engine” of the cell?
Complex I is the first, and largest, enzyme of the respiratory chain in mitochondria and bacteria. It transfers two electrons from NADH to quinone and couples (by an unknown mechanism) this process to the translocation of four protons across the membrane. Complex I plays a central role in cellular energy production, providing about 40% of the proton-motive force required for ATP synthesis. Complex I dysfunction has been implicated in many human neurodegenerative diseases. Mitochondrial complex I consists of 45 subunits, whilst the prokaryotic enzyme is simpler, consisting of 14 “core” subunits with a total mass of about 550 kDa. It represents a useful ‘minimal’ model of human complex I.
The complete structure of complex I at atomic resolution is currently unknown. Our earlier crystal structures of the hydrophilic domain of complex I from Thermus thermophilus established the electron transfer pathway from NADH through flavin mononucleotide (FMN) and seven iron-sulphur (Fe-S) clusters to the likely quinone binding site at the interface with the membrane domain. The three largest hydrophobic subunits of complex I, NuoL/Nqo12, NuoM/Nqo13 and NuoN/Nqo14 (Escherichia coli/T. thermophilus nomenclature), are homologous to each other and to the Mrp antiporter family, and so are likely to participate in proton translocation.
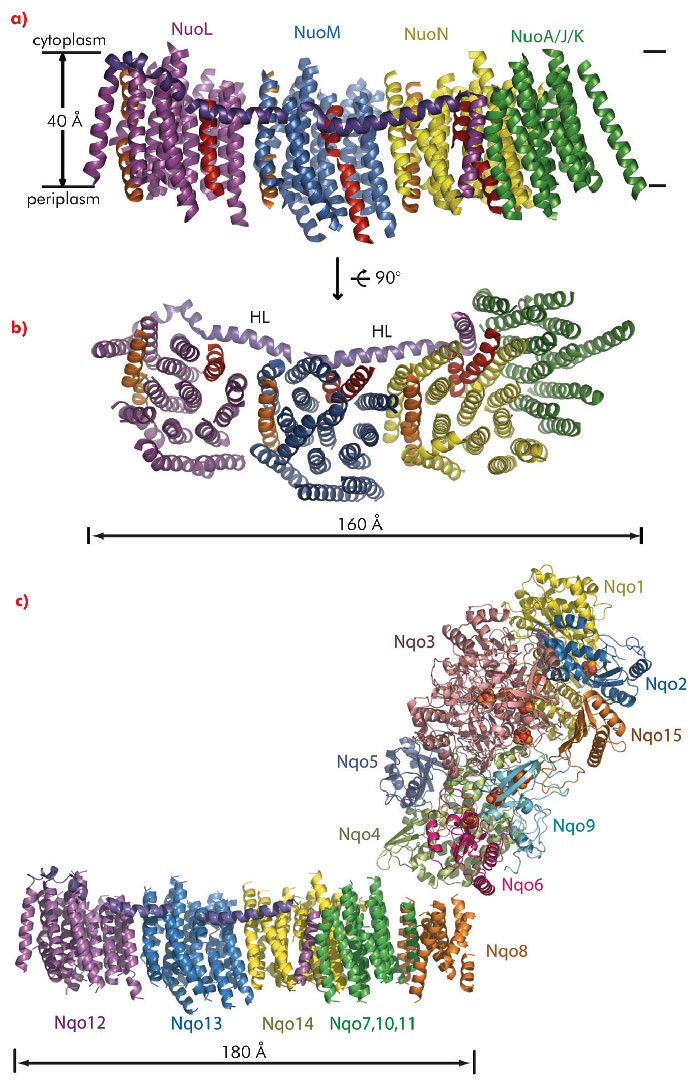
To determine the structure of the entire enzyme, we crystallised the membrane domain of complex I from E. coli. The crystals contain six subunits NuoL, M, N, A, J and K (total mass 216 kDa), and lack NuoH, which dissociates readily from the complex. After extensive optimisation, the crystals diffracted to 3.9 Å resolution, with data being collected at beamlines ID29 and ID23-2, as well as at the SLS. Phasing using TaBr cluster derivatives allowed us to build an α-helical model (Cα trace), consisting of 55 TM helices (Figure 92 a,b). A pattern of 14 TM helices repeated three times allowed us to assign these to the homologous subunits NuoL, M and N. Each subunit also contains, in the same positions, two discontinuous TM helices (shown in red and orange in Figure 92). Such helices are thought to be important for the function of many transporters and channels by introducing charge and flexibility into the membrane.
Unexpectedly, the structure revealed a 110 Å-long amphipathic α-helix, spanning almost the entire length of the membrane domain. We assigned this lateral helix (HL, purple in Figure 92) to the C-terminal extension of subunit NuoL, which is present in all species and so is likely to be functionally essential.
We also crystallised the entire complex I from T. thermophilus and obtained diffraction data to 4.5 Å resolution. The structure was solved by molecular replacement using the structure of the hydrophilic domain from T. thermophilus and the a-helical model of the membrane domain from E. coli as search probes. Subunit NuoH/Nqo8 was present in these crystals and was added to the model. The complete membrane domain now contains 63 TM helices and is one of the largest membrane protein complexes yet described (Figure 92c).
 |
|
Fig. 92: The α-helical model of the membrane domain of E. coli complex I: a) Side view, in the membrane plane; b) View from the periplasm into the membrane. c) The structure of the entire complex I from T. thermophilus. |
As seen in previous electron microscopy studies, full-length complex I is L-shaped. However, the exact positioning of the hydrophilic domain relative to the membrane domain is unexpected, as it results in a significant distance between the Fe-S cluster N2 and the predicted bilayer surface. Therefore, it appears necessary for the hydrophobic quinone to move out of the membrane by about 10 Å in order to be reduced by cluster N2. This is unusual and may be part of the coupling mechanism, aiding conformational changes.
 |
|
Fig. 93: Proposed model of proton translocation by complex I. |
The overall architecture of the complex revealed here strongly supports the idea that proton translocation is driven by long-range conformational changes (Figure 93). It is likely that movements of several helices in the hydrophilic domain (indicated by arrows in Figure 93) produce a piston-like motion of helix HL along the membrane domain. This movement can synchronously tilt the three nearby discontinuous helices, changing the conformation of ionisable residues inside the respective proton channels and resulting in the translocation of three protons. The fourth proton per catalytic cycle may be translocated at the interface of the two domains. Thus, complex I appears to resemble a steam engine, where the energy of electron transfer is used to move a piston, which then drives, instead of wheels, a set of discontinuous helices.
Principal publication and authors
R.G. Efremov, R. Baradaran and L.A. Sazanov, Nature 465, 441-445 (2010).
Medical Research Council Mitochondrial Biology Unit, Wellcome Trust/MRC Building, Cambridge (U.K.)
partners



European Synchrotron Radiation Facility - 71, avenue des Martyrs, CS 40220, 38043 Grenoble Cedex 9, France.