- Home
- Dynamics of structural transformations between the lamellar and inverse bicontinuous cubic lyotropic phases
Dynamics of structural transformations between the lamellar and inverse bicontinuous cubic lyotropic phases
Lyotropic liquid crystalline phases form when amphiphilic molecules are mixed with a polar solvent [1]. Such phases include the fluid lamellar (L![]() ; 1-D), hexagonal (HI/HII; 2-D) and inverse bicontinuous cubic phases (QIIG/QIID/QIIP; 3-D). Whilst the complex phase behaviour and energetics of such structures have received a great deal of attention over the past half century, the study of their non-equilibrium behaviour is in its infancy with little understood about the mechanisms by which one phase transforms into another. The fluid lamellar to inverse bicontinuous cubic phase transition has particular biological relevance, being structurally similar to the mechanism by which cell membranes fuse. Theoretical models for the process of membrane fusion between apposed lipid bilayers are summarised in [2]. All rely on the formation of transient lipid contacts known as stalks, which subsequently break through to form the beginnings of the tubular connections that are the fundamental connecting element in the inverse bicontinuous cubic phases. However, very few measurements have been made to test these models.
; 1-D), hexagonal (HI/HII; 2-D) and inverse bicontinuous cubic phases (QIIG/QIID/QIIP; 3-D). Whilst the complex phase behaviour and energetics of such structures have received a great deal of attention over the past half century, the study of their non-equilibrium behaviour is in its infancy with little understood about the mechanisms by which one phase transforms into another. The fluid lamellar to inverse bicontinuous cubic phase transition has particular biological relevance, being structurally similar to the mechanism by which cell membranes fuse. Theoretical models for the process of membrane fusion between apposed lipid bilayers are summarised in [2]. All rely on the formation of transient lipid contacts known as stalks, which subsequently break through to form the beginnings of the tubular connections that are the fundamental connecting element in the inverse bicontinuous cubic phases. However, very few measurements have been made to test these models.
 |
|
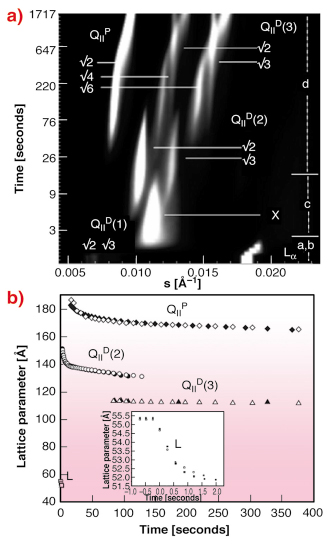
Fig. 38: The structural changes following a pressure-jump from 1100 to 260 bar, T = 46.7°C. a) each 2D diffraction image is integrated to produce a 1D plot of intensity versus scattering vector s which is then stacked as a function of time. b) shows the change in lattice parameter of all phases with time following the same pressure jump. |
The liquid crystalline lamellar (L![]() ) to double-diamond inverse bicontinuous cubic (QIID) phase transition for the amphiphile monoelaidin in excess water was effected by means of a jump in pressure and monitored using small-angle X-ray scattering (SAXS) at the high brilliance beamline ID02. Pressure jumps of up to 4 kbar can be achieved in less than 7 ms using custom-built apparatus designed and built at the University of Dortmund. We observe a remarkable sequence of structural transformations, Figure 38. A weak and previously unreported signal from a highly swollen bicontinuous cubic phase, QIID, of maximum spacing 232 Å, is consistently noted in coexistence with the fluid lamellar phase. Following the step change in thermodynamic conditions, the lamellar phase displays a marked decrease in lattice parameter. During this phase the highly-swollen bicontinuous cubic signal smears out, moves to higher scattering angle and merges with a broad, featureless ring of scatter (marked X in Figure 38). After a period of a few seconds the diffuse scatter resolves itself into a set of peaks from intermediate inverse bicontinuous cubic phases, QIID and QIIP, which replace it. Initially these intermediate cubic phases are significantly more hydrated than expected from equilibrium results. They dehydrate over time while their phase composition increases simultaneously. Such phases may be used for transport of water; the small curvature elastic energy cost associated with increasing channel size in cubic phases means this can be an energetically favourable way of increasing water flux through the sample.
) to double-diamond inverse bicontinuous cubic (QIID) phase transition for the amphiphile monoelaidin in excess water was effected by means of a jump in pressure and monitored using small-angle X-ray scattering (SAXS) at the high brilliance beamline ID02. Pressure jumps of up to 4 kbar can be achieved in less than 7 ms using custom-built apparatus designed and built at the University of Dortmund. We observe a remarkable sequence of structural transformations, Figure 38. A weak and previously unreported signal from a highly swollen bicontinuous cubic phase, QIID, of maximum spacing 232 Å, is consistently noted in coexistence with the fluid lamellar phase. Following the step change in thermodynamic conditions, the lamellar phase displays a marked decrease in lattice parameter. During this phase the highly-swollen bicontinuous cubic signal smears out, moves to higher scattering angle and merges with a broad, featureless ring of scatter (marked X in Figure 38). After a period of a few seconds the diffuse scatter resolves itself into a set of peaks from intermediate inverse bicontinuous cubic phases, QIID and QIIP, which replace it. Initially these intermediate cubic phases are significantly more hydrated than expected from equilibrium results. They dehydrate over time while their phase composition increases simultaneously. Such phases may be used for transport of water; the small curvature elastic energy cost associated with increasing channel size in cubic phases means this can be an energetically favourable way of increasing water flux through the sample.
 |
|
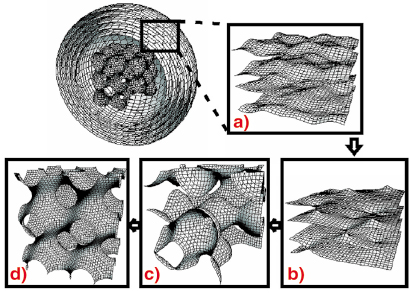
Fig. 39: The proposed structural transformations during the fluid lamellar to inverse bicontinuous cubic phase transition. |
Figure 39 illustrates the structural transformations observed in our experiments by sketching the bilayer midplane. The figures are scaled relative to each other to reflect our experimental observations. Polarising microscopy results indicate that the lamellar phase forms multi-lamellar ‘onion’ vesicles of concentric spherical bilayers. The disappearance of the highly swollen cubic phase after a thermodynamic jump coincides with the appearance of diffuse scatter. This is strong evidence that it exists inside the onion vesicle, in coexistence with the lamellar phase. Furthermore, this suggests that its function is to act as a seed for the growth of the bicontinuous cubic phases out of an initial locally-ordered bicontinuous phase (the source of the diffuse scatter). We hypothesise that the swollen cubic phase is located in the centre of the onion vesicle (Figure 39) removing the high energetic cost of filling the core with spherical bilayers of increasingly higher degrees of mechanically unfavourable curvature. Thermal undulations in the lamellar phase (a) disappear as interlamellar attachments (ILAs) are formed (b). This occurs because the bilayer’s area and the volume it circumscribes are fixed in an onion vesicle and hence forming contacts exerts a lateral tension. When the tension reaches a critical value, ILAs rupture and form a disordered network of funnels (c) through which water flows from the centre of the vesicle. The disordered sponge structure soon resolves into ordered bicontinuous cubic structures such as the swollen P cubic (d). The cubic phases, which differ little in curvature elastic energy, may then interconvert easily.
References
[1] J. M. Seddon, Biochim. Biophys. Acta 1031, 1-69 (1990).
[2] L.V. Chernomordik, M.M. Kozlov, Annu. Rev. Biochem. 72, 175-207 (2003).
Principal Publication and Authors
C.E. Conn (a), O. Ces (a), X. Mulet (a), S. Finet (b), R. Winter (c), J.M. Seddon (a) and R.H. Templer (a) Phys. Rev. Lett., 96, 108102 (2006).
(a) Department of Chemistry, Imperial College London (UK)
(b) ESRF
(c) Physical Chemistry I, University of Dortmund (Germany)
partners



European Synchrotron Radiation Facility - 71, avenue des Martyrs, CS 40220, 38043 Grenoble Cedex 9, France.