- Home
- Development of a Bacterial Biosensor for Nitrotoluenes: the Crystal Structure of the Transcriptional Regulator DntR
Development of a Bacterial Biosensor for Nitrotoluenes: the Crystal Structure of the Transcriptional Regulator DntR
The aim of this project is to develop a whole-cell bacterial biosensor for detection of nitroaromatic compounds, specifically, 2,4-dinitrotoluene (DNT). The sensor is based on a soil bacterium, Burkholderia sp. strain DNT, which is capable of using DNT as the sole source of carbon [1]. To digest this compound the bacterium produces enzymes (proteins) upon appearance of the chemical in its environment. The "device" that is used by the bacterium to detect DNT is also a protein, called a transcriptional regulator, in this case DntR. When DNT (the inducer) appears in the bacterial cell it binds to an inducer-binding cavity in DntR, which results in the initiation of reading of a DNA segment (transcription) that codes for an enzyme that is involved in the digestion of DNT. Thus, the bacterium has a feedback system, which is able to sense the compound and, as a result, synthesize enzymes that can degrade the compound. In the prototype biosensor the DNA segment coding for the enzyme was replaced by a segment coding for a green fluorescent protein. Consequently, bacteria containing this construct develop a fluorescent response in the presence of an inducer [1].
The prototype biosensor is sensitive to salicylic acid, which is an intermediate in the oxidative degradation of the aromatic ring in the bacterium. Because its sensitivity and specificity to DNT is too low, the inducer-binding cavity of DntR must be re-designed. One method to achieve this goal is to determine the three-dimensional structure of DntR and then exchange specific amino-acid residues within the cavity such that DNT would fit better.
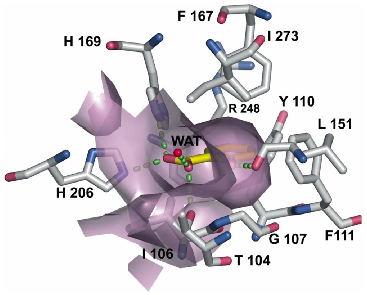
We determined two crystal structures of DntR, with acetate and thiocyanate respectively, occupying the inducer-binding cavity. In both cases DntR crystallises as a homodimer with a "head-to-tail" arrangement of monomers in the asymmetric unit. The inducer-binding domain of the DntR monomer consists of two sub-domains, each having a Rossman-fold topology. The inducer-binding cavity, with the polar entrance and the largely hydrophobic interior, is found at the interface of the sub-domains. We have positioned salicylic acid in the cavity using the positions of the acetate ions as markers. The model shows that a number of hydrogen bonds and hydrophobic interactions with the side-chains defining the surface of the cavity contribute to the specificity of the salicylate binding (Figure 79). Modelling of DNT into the inducer-binding cavity revealed several unfavourable interactions with the cavity surface and points to a number of residues that must be mutated in order to enhance the binding of DNT.
 |
|
Fig. 79: DntR containing salicylate in the inducer-binding cavities. The surface defined by protein residues is shown in purple. Carbon atoms of amino-acid residues and salicylate are coloured grey and yellow, respectively. Water molecules are shown as red spheres. Hydrogen-bond interactions are shown as green dashed lines. |
Although the asymmetric units of the crystal structures of DntR contain a homodimer, gel filtration studies demonstrate that DntR is a tetramer in solution. Examination of the packing between molecules in the crystals reveals the structure of a homotetramer in which the inducer-binding domains of DntR adopt a 'head-tail-head-tail' arrangement, similar to that recently reported for CbnR[2], which belongs to the same family of transcriptional regulators (LysR) as DntR and is the only other full-length transcription factor that has been crystallised.
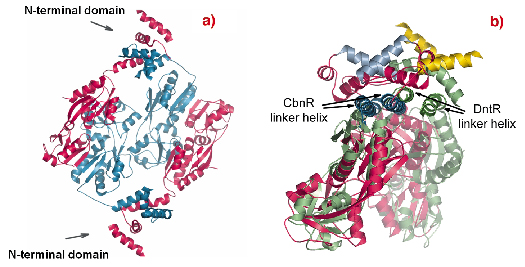
A comparison of the structures of the full-length model of DntR (Figure 80a) and that of CbnR shows a difference in the positions of their DNA-binding domains (Figure 80b). This difference is presumably due to an active (DntR) and an inactive (CbnR) conformation of the regulator, with occupied and empty inducer-binding domains, respectively. The observation is in accord with the hypothesis that transcription activation is a result of a decrease in the bend of DNA.
 |
|
Fig. 80: The structure of the full-length homotetramers. (a) DntR with the inducer-binding domain refined and the N-terminal DNA-binding domains modelled. The monomers in the head-to-head dimers are shown in red and blue. (b) A superposition of the DntR dimer (green ribbons with recognition helices coloured yellow) and the CbnR dimer (red ribbons with recognition helices coloured blue). |
In conclusion, we have solved the structure of DntR, which paves the way for the design of a bacterial biosensor for nitrotoluenes. In addition, the structure provides insights into the mechanism of transcription regulation by the LysR regulatory proteins.
References
[1] L.C. Ng, M. Forsman, FOI Research Report (2000) (http://www.foi.se/).
[2] S. Muraoka, R. Okumura, N. Ogawa, T. Nonaka, K. Miyashita, T. Senda, J. Mol. Biol. 328, 555-66 (2003).
Principal Publication and Authors
I.A. Smirnova (a), C. Dian (a,b), G.A. Leonard (b), S. McSweeney (b), D. Birse (a,c), P. Brzezinski (a), J. Mol.Biol. 340, 405-418 (2004).
(a) Department of Biochemistry and Biophysics, Stockholm University (Sweden)
(b) ESRF
(c) TGN Biotech Inc., Québec (Canada)
partners



European Synchrotron Radiation Facility - 71, avenue des Martyrs, CS 40220, 38043 Grenoble Cedex 9, France.