- Home
- Structure of a Hepatitis C Virus Immunodominant Antigenic Site
Structure of a Hepatitis C Virus Immunodominant Antigenic Site
Hepatitis C virus (HCV) is the major causative agent of transfusion associated hepatitis. Among the structural proteins of the virus, the core protein is highly conserved in the various HCV genotypes and elicits a rapid response after the onset of the disease. Thus, the measurement of HCV antibodies (Abs) titer in serum is widely used to screen for HCV infection [1].
Several studies, mainly based on peptide scanning using chemical or recombinant approaches, have established that the first 120 residues of the HCV core protein contain highly immunogenic B cell epitopes that are predominantly recognised by HCV-infected patient serum; one of them has been identified in the N-terminal region (20-45) of the core protein.
To analyse the structural characteristics of that HCV-immunodominant epitope we generated different monoclonal antibodies (mAbs) by immunising mice with a truncated recombinant 1-120 region. Among the mAbs obtained, 19D9D6 appeared to be specific for an epitope encompassed by residues 29-33 [2]. This antibody was crystallised as was its complex with a synthetic 28 amino-acids peptide that encompasses the recognised portion 13-40 of core. Crystals of the free antibody (P21212; a = 63.48 Å, b = 173.68 Å, c = 41.73 Å) diffracted to 1.6 Å resolution on beamline ID14-1 while crystals of the complex (P21; a = 42.18 Å, ß = 101.7 Å, c = 55.18 Å, ß = 98.6°) diffracted to 2.34 Å resolution on beamline BM30.
Both structures were solved by molecular replacement followed by rigid body refinement and cycles of conjugate gradient and isotropic temperature factor refinement using CNS.
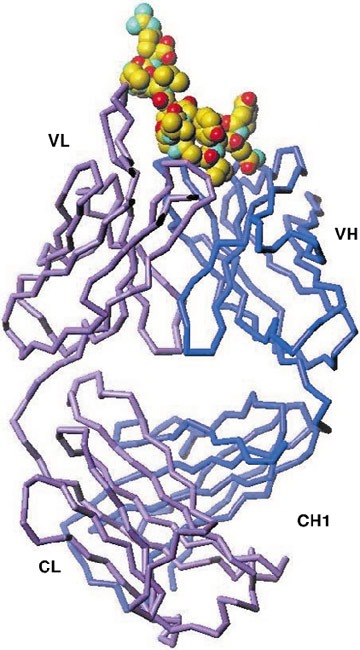
The final electron density maps of the complex between the 19D9D6 Fab' and the 13-40 peptide allows the positioning of all the 438 residues of the Fab' and 15 of the 28 peptide residues. The peptide is bound in a crevice between the VL and the VH domains (Figure 26). It was a curved construction and only nine residues interact with mAb 19D9D6. Detailed comparison of the free and complexed 19D9D6 Fab' structures shows that only minor structural adjustments are needed in the VL domain to accommodate binding of the peptide. The crystal and solution structures of the core peptide are different. In solution, its small hydrophobic core is protected by two helices that pack against each other. In the crystal, the peptide conformation is stabilised by hydrogen bonds and van der Waals interactions with the antibody. Only a short stretch of five residues can be superimposed between the solution and crystal structures.
 |
|
Fig. 26: Schematic representation of the complex of core peptide (RNTNRRPQDVKFPGGGQIVGGVYLLPRR) and 19D9D6 antibody. |
This is the first structure of a fragment of HCV core antigen. The structure of the complex of Fab' 19D9D6 with core peptide 13-40 offers an unique opportunity to understand how antibodies act in acqueous solutions and in lipidic environments. Given that this monoclonal antibody competes effectively with human sera, it is suggested that human anti-HCV Abs may share many of the features of mAb 19D9D6, including the ability to destabilise, partially at least, the solution conformation of the antigen often found associated with a lipidic phase.
References
[1] H.J. Park, S.M. Byun, Y.J. Ha, J.S. Ahn and H.M. Moo, J. Immunoassay, 16, 167-181 (1995).
[2] C. Jolivet-Reynaud, P. Dalbon, F. Viola, S. Yvon, G. Paranhos-Baccala, N. Piga, L. Bridon, M.A. Trabaud, N. Battail, G. Sibai and M. Jolivet, J. Med. Virol., 56, 300-309 (1998).
Principal Publication and Authors
R. Ménez (a), M. Bossus (a), B.H. Muller (a), G. Sibai (b), P. Dalbon (b), F. Ducancel (a), C. Jolivet-Reynaud (c) and E.A. Stura (a), The Journal of Immunology, 170, 1917-1924 (2003).
(a) UMR CEA-bioMérieux DIEP, Saclay (France)
(b) R&D bioMérieux, Marcy l'Etoile (France)
(c) UMR CNRS-bioMérieux, Lyon (France)
partners



European Synchrotron Radiation Facility - 71, avenue des Martyrs, CS 40220, 38043 Grenoble Cedex 9, France.