- Home
- Users & Science
- Scientific Documentation
- ESRF Highlights
- ESRF Highlights 2013
- Structural biology
- Structural basis for stabilisation of Hsp70-substrate complexes by Hip
Structural basis for stabilisation of Hsp70-substrate complexes by Hip
Under laboratory conditions, many proteins fold spontaneously into their biologically active conformation. In the crowded cytosol of the cell, however, larger proteins in particular have a tendency to mis-fold and aggregate, a process that can disrupt vital cellular processes. Cells therefore strive to maintain ‘proteostasis’, i.e. a healthy proteome, by carefully controlling synthesis and folding of new proteins as well as the degradation of those that are damaged. Failure to do this may lead to the accumulation of potentially toxic, mis-folded protein species and may cause severe conditions such as Alzheimer’s or Parkinson’s disease. A principal component of the cellular proteostasis machinery, a network of multiple factors, is a set of proteins known as molecular chaperones, which function to prevent protein aggregation and ensure that folding occurs on a physiologically relevant time scale [1].
The highly-conserved molecular chaperone Hsp70 is a central component of the protein folding machinery in bacterial and eukaryotic cells, both interacting with nascent polypeptides on translating ribosomes and cooperating with chaperone factors and protein degradation machineries that act down-stream. Hsp70 functions as a molecular machine, using ATP hydrolysis to cycle between conformational states with distinct binding properties for non-native polypeptides [2]. Tight interaction with hydrophobic peptide motifs occurs in the ADP-bound state while the binding of ATP triggers substrate release. Because Hsp70 has a low intrinsic ATP hydrolysis activity with slow ADP dissociation, it is critically dependent on so-called cochaperones, which stimulate ATP hydrolysis and accelerate ADP-ATP replacement, J-domain proteins and nucleotide exchange factors, respectively. A plethora of these cofactors, occurring specifically in eukaryotic cells, serve to recruit Hsp70 for specific functions, such as the disassembly of clathrin coats around vesicles after endocytosis. In addition to J-domain proteins and exchange factors, metazoans, plants and numerous protists contain yet another cytosolic Hsp70-co-chaperone named Hip (Hsc70-interacting protein), which stabilises Hsp70-substrate complexes. Together with Hsp70, Hip was implicated in the prevention of aggregation of α-synuclein and the poly-Q androgen receptor as well as in the timely degradation of hyper-phosphorylated Tau. All of these proteins form deposits in neurodegenerative diseases [3,4].
 |
|
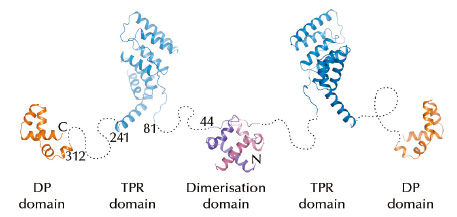
Fig. 54: A tentative model for the structure of a complete Hip dimer. The flexible linkers (dashed lines) are not drawn to scale. |
Using SAXS and diffraction data collected at the ESRF, we showed that Hip has a modular domain architecture consisting of an N-terminal dimerisation domain, a central TPR module and a C-terminal DP domain that are connected by flexible linkers (Figure 54). A crystal structure of 1.1 Å resolution revealed that the N-terminal dimerisation domain, comprising only 44 residues from each chain, forms a unique compact helix bundle fold. The TPR domain of Hip is sufficient to mediate the interaction with Hsp70 and, in contrast to other TPR domain cofactors of Hsp70, recognises the nucleotide binding domain (NBD) of Hsp70 and not the C-terminal region of the chaperone.
 |
|
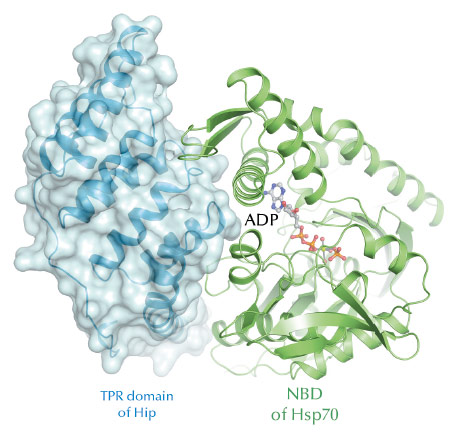
Fig. 55: The crystal structure of the Hip-Hsp70 core complex. ADP and phosphate (shown in stick representation) are locked in the Hsp70 nucleotide binding cleft (NBD). |
Biochemical assays showed that ADP dissociation from Hsp70 is slowed by complex formation with Hip. The crystal structure of the core complex of Hip and Hsp70, consisting of the TPR domain of Hip and the NBD of Hsp70, determined with data from beamline ID23-2, revealed the structural basis for this interaction (Figure 55). In the complex, Hip acts as a molecular clamp, locking ADP in the nucleotide binding cleft of Hsp70, and occludes the common interaction site for the eukaryotic Hsp70 nucleotide exchange factors, thus blocking active cycling of the molecular chaperone. However, a biologically relevant affinity is reached only when Hip specifically recognises a substrate with its C-terminal domain, or when both arms of the Hip dimer are engaged in interaction with Hsp70, allowing normal processing of bona-fide Hsp70 substrates. This mechanism explains how Hip enhances aggregation prevention by Hsp70 and facilitates the transfer of substrate proteins to down-stream chaperones or the proteasome.
Principal publication and authors
Z. Li, F.-U. Hartl and A. Bracher, Nature Structural & Molecular Biology 20, 929-935 (2013).
Department of Cellular Biochemistry, Max-Planck-Institute of Biochemistry, Martinsried (Germany)
References
[1] Y.E. Kim et al., Annu Rev Biochem 82, 323-355 (2013).
[2] M.P. Mayer, Mol Cell 39, 321-31 (2010).
[3] C. Roodveldt et al., EMBO J 28, 3758-3770 (2009).
[4] A.M. Wang et al., Nat Chem Biol 9, 112-118 (2013).
partners



European Synchrotron Radiation Facility - 71, avenue des Martyrs, CS 40220, 38043 Grenoble Cedex 9, France.