- Home
- Users & Science
- Scientific Documentation
- ESRF Highlights
- ESRF Highlights 2001
- Life Sciences
- Structure of a Membrane Protein Complex: Formate Dehydrogenase-N at 1.6 Å
Structure of a Membrane Protein Complex: Formate Dehydrogenase-N at 1.6 Å
The respiration of nitrate constitutes a major respiratory pathway in Escherichia coli under anaerobic conditions. A major electron donor in this pathway is formate, which is produced from pyruvate via acetyl-CoA. A system composed of the integral membrane proteins, formate dehydrogenase-N (Fdh-N) and dissimilatory nitrate reductase (Nar) utilises the two-electron oxidation of formate as an electron donor for the reduction of nitrate to nitrite. Nar/Fdh-N has a redox loop mechanism responsible for this energy conservation. The Nar/Fdh-N system generates proton motive force, which is used by ATP synthase and secondary transporters, by the redox loop mechanism, a mechanism ubiquitous among biomembranes of higher organisms and bacteria.
Both Fdh-N and Nar are members of a subgroup of molybdo-enzymes, binding the molybdopterin guanine dinucleotide (MGD) form of the molybdopterin cofactor in their active site. Both enzymes are three-subunit proteins (![]() , ß,
, ß, ![]() ), consisting of two membrane-associated subunits and an integral membrane subunit. The crystal structure of Fdh-N was determined by multiple anomalous dispersion (MAD) using the 22 native Fe atoms. MAD and high-resolution data sets were collected at beamlines ID29 and ID14-3, respectively. Automated model building in combination with phase extension to 1.6 Å was performed using the ARP/wARP program suite. Refinement, including water molecule placement, was performed using the programs ARP and CNS with final Rcryst of 17.7 % and Rfree of 19.5 %. This is the highest resolution structure achieved for any membrane protein complex to date (Figure 1).
), consisting of two membrane-associated subunits and an integral membrane subunit. The crystal structure of Fdh-N was determined by multiple anomalous dispersion (MAD) using the 22 native Fe atoms. MAD and high-resolution data sets were collected at beamlines ID29 and ID14-3, respectively. Automated model building in combination with phase extension to 1.6 Å was performed using the ARP/wARP program suite. Refinement, including water molecule placement, was performed using the programs ARP and CNS with final Rcryst of 17.7 % and Rfree of 19.5 %. This is the highest resolution structure achieved for any membrane protein complex to date (Figure 1).
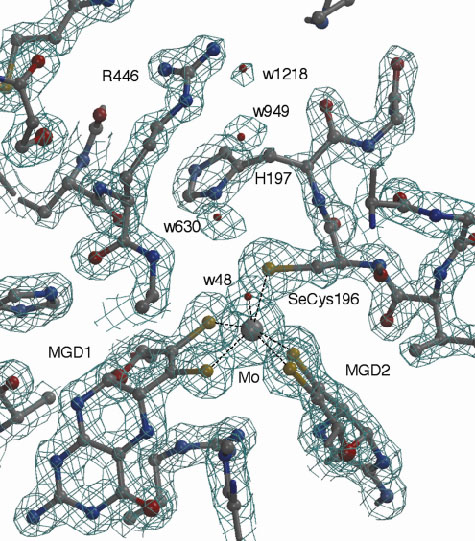 |
Fig. 1: Electron density map around the active site of Fdh-N at 1.6 Å resolution. |
The overall structure of Fdh-N is shown in Figure 2. Fdh-N is packed as a trimer (total MW 510 kDa) with the monomers related by a crystallographic threefold symmetry axis. The trimer shows a "mushroom"-like shape with the largest dimensions of 125 Å (along the membrane) x 150 Å (along the membrane normal). The ![]() -subunit incorporates the catalytic domain with a molybdenum (Mo) atom, two MGD cofactors, a selenocysteine residue and one [4Fe-4S] iron-sulphur cluster. The ß-subunit is an electron transfer unit containing four [4Fe-4S] iron-sulphur clusters, while the membrane intrinsic
-subunit incorporates the catalytic domain with a molybdenum (Mo) atom, two MGD cofactors, a selenocysteine residue and one [4Fe-4S] iron-sulphur cluster. The ß-subunit is an electron transfer unit containing four [4Fe-4S] iron-sulphur clusters, while the membrane intrinsic ![]() -subunit incorporates the two heme b molecules and a quinone reduction site. The structure shows how electrons are transferred from formate to MQ through MGD, 5 [4Fe-4S] clusters and two heme b groups, a total distance of over 90 Å. Further studies utilising a menasemiquinone analogue, HQNO (2-n-nonyl-4-hydroxyquinoline N-oxide) revealed the MQ binding site in the g-subunit and a possible proton uptake pathway from the cytoplasm to this quinone binding site. A comparative study of the Fdh-N with the related enzymes, nitrate reductase and [NiFe] hydrogenase, has successfully explained how the proton motive force is generated by the Fdh-N/Nar system.
-subunit incorporates the two heme b molecules and a quinone reduction site. The structure shows how electrons are transferred from formate to MQ through MGD, 5 [4Fe-4S] clusters and two heme b groups, a total distance of over 90 Å. Further studies utilising a menasemiquinone analogue, HQNO (2-n-nonyl-4-hydroxyquinoline N-oxide) revealed the MQ binding site in the g-subunit and a possible proton uptake pathway from the cytoplasm to this quinone binding site. A comparative study of the Fdh-N with the related enzymes, nitrate reductase and [NiFe] hydrogenase, has successfully explained how the proton motive force is generated by the Fdh-N/Nar system.
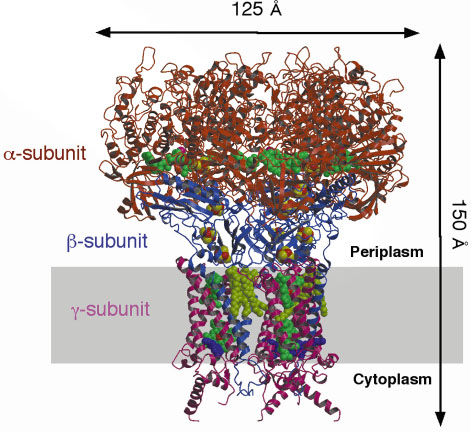 |
Fig. 2: Trimer of Fdh-N viewed parallel to the membrane. Catalytic |
Principal Publication and Authors
M. Jormakka (a), S. Tornroth (c), B. Byrne (b), S. Iwata (a, b, c) and A. Thompson (d), Science, in press (2002).
(a) Division of Biomedical Sciences, Imperial College (UK)
(b) Department of Biological Sciences, Imperial College (UK)
(c) Department of Biochemistry, Uppsala University (Sweden)
(d) EMBL Grenoble Outstation (France)
partners



European Synchrotron Radiation Facility - 71, avenue des Martyrs, CS 40220, 38043 Grenoble Cedex 9, France.